
IV. L'oreille interne

1. Le fonctionnement biologique précis de la cochlée.
C'est maintenant que nous approchons de plus près les mécanismes complexes qui constituent notre capacité à interpréter les variations de pression que sont le son dans notre environnement.
A. La cochlée, première approche
1. Rappels
La cochlée , organe clé de l'audition situé dans l'oreille interne , à proximité des organes de l'équilibre (système vestibulaire) , prend la forme d'une structure osseuse en forme de coquille d'escargot.
Rappel de l'emplacement de la cochlée dans l'oreille interne , en vue externe :
Le vestibule et les canaux semi-circulaires sont les organes de l'équilibre, nous ne nous y intéressons pas ici
Sur l'image ci-dessus, on peut voir le vestibule (en bleu) ainsi que la cochlée et son apex (l'extrémité "pointe" de la cochlée). Il est intéressant d'observer que les nerfs de la cochlée et du vestibule se rejoignent , pour former le VIIIème nerf crânien.

2. Introduction a l'organisation de la cochlée

La cochlée est en réalité constituée d'un tube osseux creux de forme conique, appelé canal spiral , mesurant environ 35 mm de long , ainsi que de son contenu. Le canal spiral est enroulé en deux tours autour d'un axe creux lui aussi, le modiolus. Le premier tour est appelé tour basal , le second est appelé tour apical (car proche de l'apex). Le modiolus renferme le point de départ du nerf auditif.
"L'enveloppe" osseuse externe de la cochlée s'appelle la capsule osseuse.
Représentation de la cochlée en coupe:
A l'intérieur du canal spiral, qui est, nous l'avons dit, creux, la cochlée n'est pas un espace uni : en effet elle est constituée de trois canalicules.
Ces trois canalicules sont :
-La rampe tympanique
-La rampe vestibulaire
-Le canal cochléaire
Pour visualiser :

Tous les éléments décrits ici s'étendent le long de tout le canal spiral , et sont toujours disposés de la même façon (excepté à l'apex , point culminant de la cochlée).
La lame spirale osseuse est une petite excroissance osseuse aplatie de forme hélicoïdale qui sépare partiellement le canal spiral en deux de façon horizontale, avec d'un coté la rampe tympanique , et de
l'autre le canal cochléaire de forme triangulaire ainsi que la rampe vestibulaire.
C'est la membrane basilaire qui achève de scinder le canal.
La rampe tympanique et la rampe vestibulaire sont remplies d'un liquide :
le périlymphe. Elles sont globalement étanches, excepté en un point où elles se
rejoignent par un petit orifice, à l'apex : l'hélicotrème.
Le canal cochléaire est lui aussi rempli d'un liquide : l'endolymphe.
Le canal cochléaire se situe entre la rampe tympanique et l' Organe de Corti ,
qui est l'organe transformant les ondes sonores amenées à la cochlée, en signaux
nerveux envoyés ensuite au cerveau. Ces signaux passent par le ganglion spiral
(ou ganglion cochléarien), contenu dans la lame spirale et relié au nerf auditif,
dans le modiolus.
L'Organe de Corti repose sur la membrane basilaire , qui se trouve donc entre la rampe tympanique et l'Organe de Corti.
La membrane basillaire est l'organe qui transmet l'onde sonore dans toute la longueur de la cochlée à l'Organe de Corti. C'est pourquoi il est nécessaire de comprendre que l'organe de Corti repose sur cette membrane.
Voici un schéma de coupe transversale du canal spiral , représentant les différents éléments présentés ci-dessus de façon plus claire :

B. Les canalicules ou l'organisation spatiale générale
des éléments constitutifs de la cochlée
1. Introduction à la membrane basilaire
2. Un peu d'histoire
Comme nous l'avons expliqué plus tôt , la chaîne des osselets fait passer la vibration acoustique du milieu aérien de l'oreille externe aux liquides contenus par la cochlée. Cela entraîne une vibration/oscillation générale du contenu du canal spiral.
La membrane basillaire est le vecteur permettant de répartir dans la cochlée les différentes fréquences sonores afin de les analyser.
En effet la membrane basilaire va , lorsque le son arrive dans la cochlée , vibrer.
Lorsque la vibration acoustique est transmise à la membrane basilaire , cette dernière décrit une onde.
L'onde en question se propage dans la membrane en augmentant d'amplitude jusqu'en un point à partir duquel elle disparaît rapidement. Ainsi le point culminant de l'onde, point où elle est codée, dépend de la fréquence du son.
Lorsque la cochlée reçoit un son grave, ayant donc une fréquence faible , la membrane basilaire transmettre la vibration jusqu'en un point proche de l'apex.
Tandis que lorsque le son est aigu , avec donc une fréquence élevée , la membrane basilaire va transmettre la vibration jusqu'en un point plus proche de l'entrée de la cochlée (fenêtre ovale décrite en partie ) .
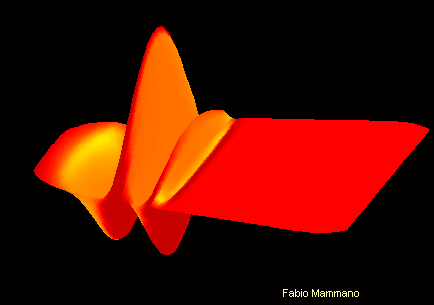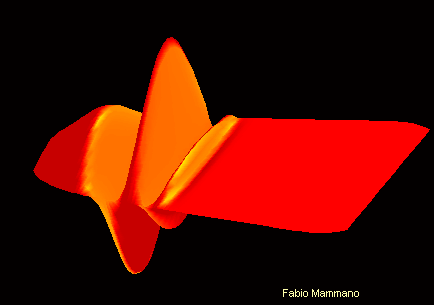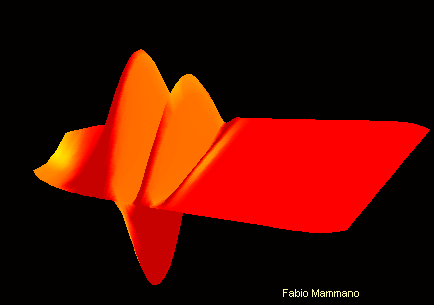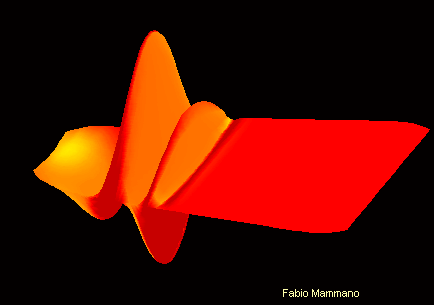
Pour comprendre :
Son de fréquence 400Hz sur la membrane basilaire Son de fréquence 4kHz sur la membrane basilaire
On peut ici visualiser le mouvement différencié de la membrane basilaire en fonction de la fréquence sonore. L'apex se trouve ici vers la droite de l'image , et la fenêtre ovale vers la gauche.
Cette propriété est en partie due à une diversité dans les propritétés physiques de la membrane basilaire, qui est moins large et plus rigide à la base de la cochlée qu'à l'apex.
La compréhension du fonctionnement de la membrane basilaire et de la logique de répartition des sons pour le codage nerveux s'est faite progressivement.
La première hypothèse décisive fut formulée par le physicien et physiologiste allemand Hermann von Helmholtz (1821-1894) en 1885.
Selon lui le codage différentié des sons se faisait selon l'emplacement le long de la membrane basilaire. Il supposait que la membrane était constituée de petits « fragments » ayant une réponse physique à une fréquence particulière.
Selon ce modèle donc, les cellules de la membrane basilaire sont sensibles à une vibration spécifique , mais le message nerveux transmis par l'Organe de Corti au niveau de chaque partie de la membrane est identique ; il est différencié par le système nerveux qui en fonction de sa provenance reconnaît la fréquence sonore correspondant.
Cette théorie fut invalidée que 63 ans plus tard , en 1948 , lorsque Georg von Bekesy (1899-1972) , biophysicien hongrois , démontra que ces « fragments » n'existaient pas et établit un nouveau modèle : le codage du son dépendrait selon lui de la distance parcourue par l'onde dans la membrane basilaire.
La répartition de la vibration le long de la membrane n'est selon lui pas due à un phénomène cellulaire spécifique mais aux propriétés physiques de cette membrane (moins large et plus rigide à la base de la cochlée qu'à l'apex). Un son de haute fréquence dépensera toute son énergie à la base de la membrane et l'onde ira peu avant dans la cochlée ; un son de basse fréquence se propagera le long de la membrane plus loin vers l'apex.
Ainsi selon le modèle de Georg von Bekesy , lorsque un point spécifique de la membrane basilaire vibre avec une certaine fréquence (par exemple 1kHz) , l'Organe de Corti les décode et envoie des impulsions nerveuses avec cette même fréquence (ici donc 1000 impulsions par seconde).
Or ce modèle pose un problème : il est globalement admis aujourd'hui que la cochlée ne réalise pas d'analyse du son ; elle transmet directement les informations , organisées , au cerveau qui se charge de les analyser.
Avec le modèle de Bekesky , les neurones devraient transmettre l'information relative aux fréquences très élevées , à partir de 4000-5000Hz, en émettant des impulsions nerveuses avec la même fréquence. Seulement les neurones ne peuvent décharger des impulsions trop rapidement. Tous les sons ne seraient donc pas audibles.
Dans les faits, les deux différents modèles susdécrits sont en partie vrais. La répartition et le codage des sons dans la membrane basilaire et l'Organe de Corti est faite selon un modèle de fréquence et de place.
En effet pour les fréquences basses , le processus de répartition et le codage est fréquentiel , comme dans le modèle de Bekesy.
Pour les hautes fréquences (à partir de 4000-5000Hz) en revanche, le processus suit le principe de fonctionnement du modèle de Helmholtz (tout en étant différent au niveau cellulaire ; cela est très complexe et nous ne nous étendrons pas dessus), puisque comme nous l'avons dit les neurones ne pourraient pas suivre un système fréquentiel.
De plus il a été démontré par Bekesy également que les cellules codant l'information le font pour un fréquence spécifique, mais qu'elles le font aussi de façon moins importante pour les fréquences voisines. Cela permet de déconstruire et coder les sons complexes en reconnaissant la fréquence fondamentale et les harmoniques.
Répartition du codage des fréquences sonores le long de la cochlée (tonotopie) :
cliquez pour zoomer
pitch = hauteur
C. La membrane basilaire


3. Une répartition mixte
